日本漁業種苗放流近一甲子的回顧
2018/03/13
示意圖

日本漁業種苗放流近一甲子的回顧
臺灣國立海洋大學水產養殖系
郭金泉教授 整理
全世界實施水產生物放流的國家以日本放流的種類與數量最多,包含魚類、水產無脊椎動物(貝類、海膽、海蔘、甲殼類蝦蟹等)。二戰後日本經濟自 1960 年代快速復興,鑒於土地不足乃填海造陸,工廠林立,吸引無數農漁村人口流入成為工廠勞工,人口持續往大城市集中。為了彌補因為填海造陸造成漁業環境之破壞與沿岸漁業資源枯竭的現象,同時也企圖挽留因為經濟發展而外流的漁村子弟,日本政府 1963 年在瀨戶內海開始栽培漁業,實施種苗放流,希望可以增加沿岸漁業資源。筆者獲科技部贊助邀請日本國東京海洋大學北田修一教授蒞臺演講日本漁業種苗放流並與臺灣官學界交流。本文是筆者綜合北田修一教授於 2016 年 11 月 23~24 日兩天在國立臺灣海洋大學,行政院農委會水產試驗所基隆總所、農委會漁業署以及國立臺灣大學漁業科學所之演講內容整理而成,謹共饗臺灣讀者。

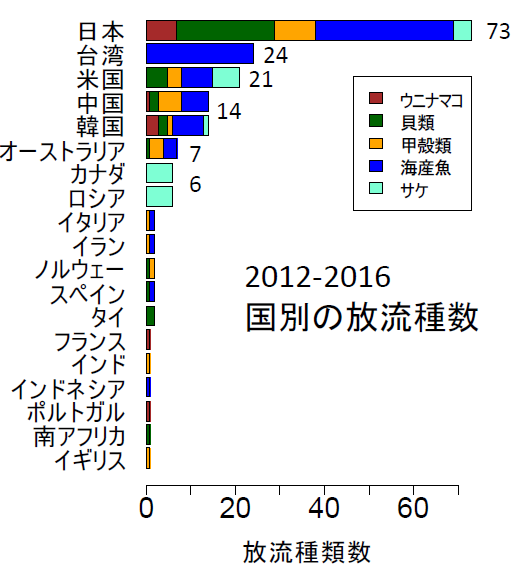
北田修一教授整理 2012 至 2016 全球放流水產種苗物種數的國家,台灣放流 24 個物種,主要是魚類、僅次於日本,是世界放流水產種苗物種第二多的國家。
那到底放流水產生物種苗有沒有效?對野生族群的生態與遺傳構造會不會產生影響?首先,研究者必須開發出有效標記生物的方法,才能正確判斷捕獲的水產生物是野生或是養殖放流生物的再捕獲,也方可執行後續的種苗放流效果評估。

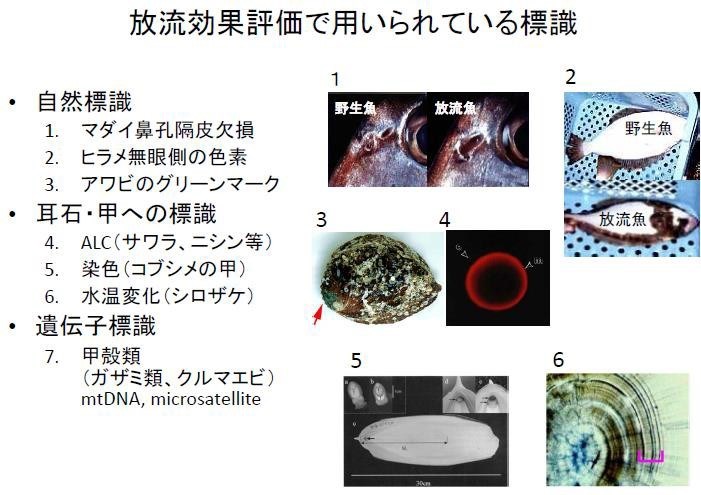
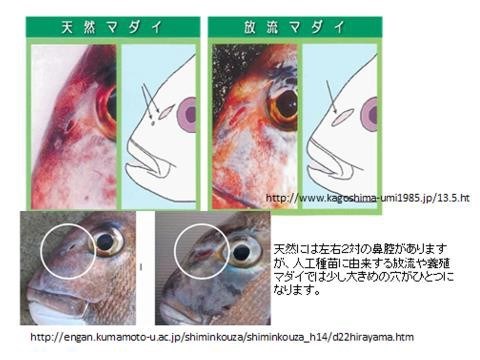
目前日本使用數種水產生物的標記方法:(1) 自然標記法:如真鯛(嘉鱲)密集養殖多代後,兩鼻孔(入水孔和出水孔)會變成僅一鼻孔;養殖的比目魚,沒有眼睛白色部位的體側會出現黑斑色素;養殖九孔與鮑魚依初期餵食餌料之不同,會在貝殼的基點留下餵食餌料之色素,因為養殖和野生鮑魚餌料差異,研究人員可以殼色辨別是養殖放流的鮑魚或是野生鮑魚。(2) 剪掉或拔除身體某部位:水產無脊椎動物藉由剪除身體某部位,如螃蟹的游泳肢、蝦子之剪尾,不但標示作業費心, 勞力密集,且有些部位仍會再生長回,辨別度不高,死亡率也大。(3) 染色生物硬組織:如魚類耳石、烏賊骨板的 ALC (Alizarin red) 染色,不過現在認為 ALC 會衍生食害問題已被禁用。(4) 遺傳標記:因費用較高,尚未普及。

放流效果評估的方法
日本在 1980 年代以前是在放流生物體外打上標記(tag)再放流,捕獲到此有標記生物的民眾再回報研究單位,但是由於回報率很低,可信度不高效果不彰;後來改採統計的採樣理論,派調查員到漁市場檢查市場當天總漁獲的每個個體,以自然標記法判斷魚隻是養殖放流的或是野生的魚隻,就非常準確。回收報告率趨近百分百。

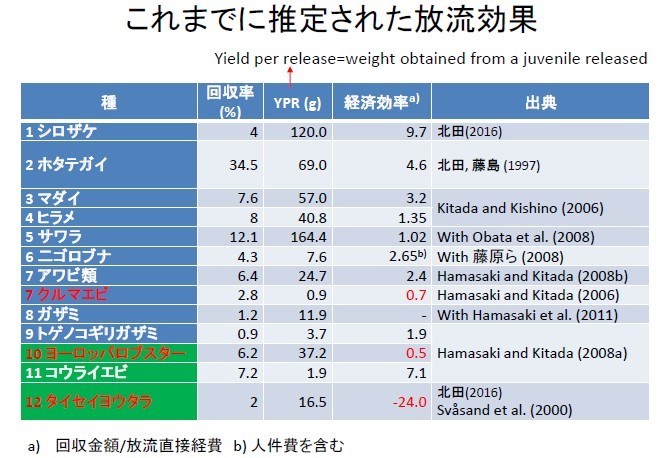
根據此可靠的回收報告數據,日本估算出回收率(%)、YPR(g:yield per release:放流每尾仔魚的重量回收)、及經濟效益。根據北田所作放流的經濟效益評估研究結論,只有少數(鮭魚、帆立貝、真鯛)幾種放流例子具經濟效益,斑節蝦(クルマエビ)的放流是賠本事業。挪威甚至還因不合成本(表上紅字),放棄龍蝦(ロブスタ-)與鱈魚(タイセイヨウタラ)的放流事業,而全力發展大西洋鮭魚養殖。
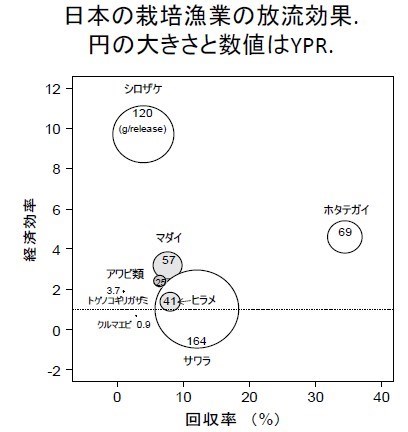
圓圈表示 YPR(yield per release:放流每尾仔魚的重量回收),越大表示此種生物長得快。例如放一尾鰆魚(サワラ)仔魚的 YPR 可以回收到 164 克、而且回收率約 12%, 經濟效益划算。所以放流生物長得越快(YPR)、而且活存率(回收率)越好、具高經濟價值的水產生物,放流效益較好、方可以有效回收放流成本。目前大部分水產生物的放流效益都是政府賠本在經營。虛線(= 1)代表損益平衡。
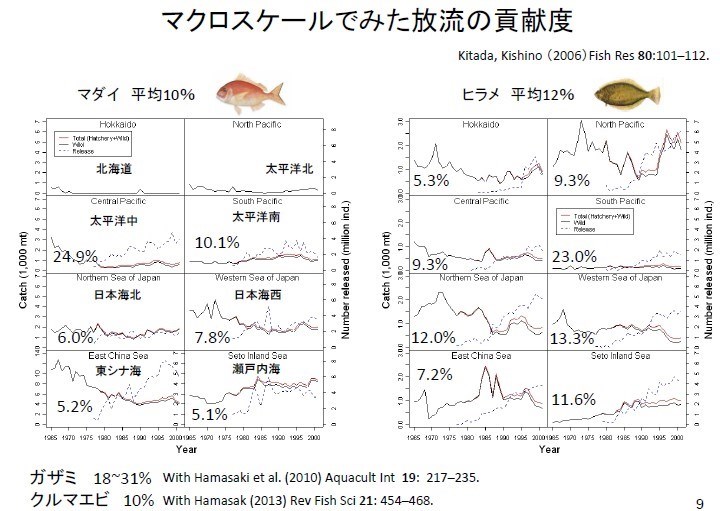
日本在不同海區大規模放流真鯛(マダイ),紅線表總捕獲量(養殖放流量+野生量)、虛線表放流養殖真鯛的數量、黑線表以 YPR 換算放流養殖真鯛數量的捕獲量。放流養殖真鯛數量的貢獻就是紅線與黑線的差,依各個海域放流養殖真鯛數量的貢獻度由 5.1~24.9%不等,平均嘉鱲的放流效果大概是 10%。日本北海道和太平洋北沒放流真鯛。比目魚(ヒラメ)貢獻度大概是 12%,螃蟹(カザミ)貢獻度介於 18 和 31%之間,斑節蝦(クルマエビ) 貢獻度更小僅有10%。可能放流量遠比自然資源的變動量(入添量)小,種苗放流效果很難由自然資源的變動量分離出來、所以放流直接效果(貢獻度)並不明顯。
1974年~鹿兒島的真鯛(Pagrosomus major,嘉鱲)放流栽培漁業
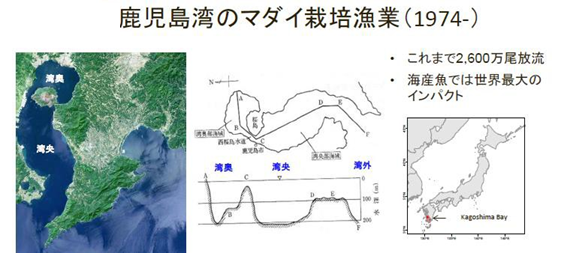
1970 年以後日本國九州鹿兒島灣真鯛的魚獲量急遽下降,日本官方於 1970 年代後期開始大量放流真鯛魚苗。據統計鹿兒島灣自 1974 以來日本官方總共放流 2600萬尾嘉臘魚,是世界上最大的海水魚放流量,對生態衝擊很大。追蹤 1970 至 2012 年鹿兒島灣真鯛總漁獲量(藍線),野生魚捕獲量(綠線),養殖漁獲量(紅線),魚苗放流量(紅虛線)的經年變動,發現隨著魚苗放流量減少(1990 年代至 2000 年),此時期總漁獲量(藍線)也減少。2005 年以後即使放流魚苗數減少,野生魚捕獲量(綠線)卻增加。
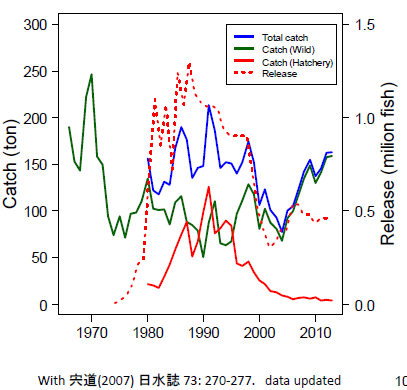
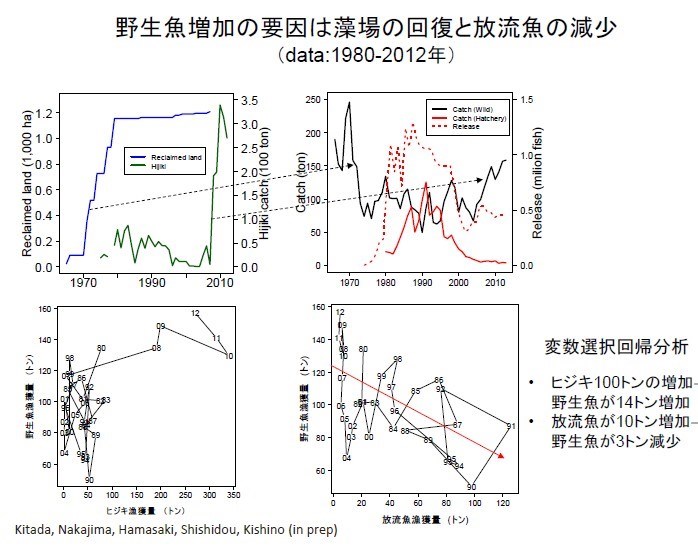
解析其原因發現:日本 60 年代經濟起飛,1970 年以後由於鹿兒島灣棲地遭大規模破壞(左上圖藍線上揚),真鯛數量(右上圖黑線陡降)急速減少。但是為何鹿兒島灣裡面的真鯛魚獲量(右上圖黑線)會在 2009 年後增加(右上圖黑線上攀),主要的原因是鹿尾菜海藻(ヒジキ;左上圖綠線急增)的回復;而且鹿尾菜海藻的回復必須達某一定量(50 公噸;左下圖)以上,才會發生幫助真鯛數量增加的效應(2009 年綠線;右上圖)。如果放流越多養殖真鯛魚苗,野生真鯛的捕獲量就越少(右下圖);但是如果每增加一百公噸鹿尾菜海藻,野生真鯛就會增加 14 公噸,因為鹿尾菜海藻也可以提供真鯛的棲地。如果放流 10 公噸養殖真鯛魚苗,野生真鯛漁獲反而會減少3 公噸,因為鹿兒島灣真鯛的生物承載量是一定的。

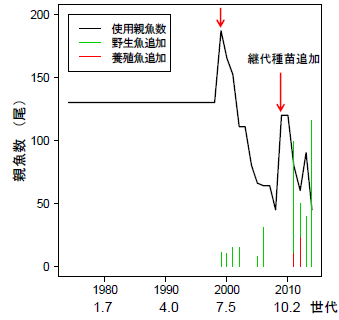
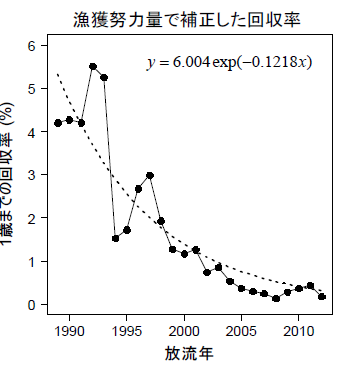
由於使用的真鯛種魚是 1980 年 130 尾種魚繼代培養子孫之種魚,種魚培養出來的放流魚隻的回收率有逐年遞減現象;研究指出每一個世代其活(生)存率減少約12%。
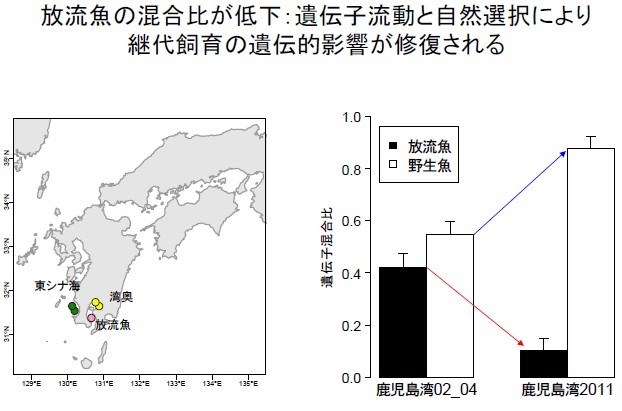
鹿兒島灣內部(灣奧;黃圓圈),2004 年養殖真鯛魚的基因混合比(黑長條)一開始佔比蠻高,和野生魚相差不多;但是隨著時間拉長,佔比漸漸變低;野生魚(白長條) 佔比在 2011 年已遙遙領先養殖魚的基因(黑長條)。此乃鹿兒島灣內部(灣奧; 黃圓圈)和鹿兒島灣外部(東海;綠圓圈)的真鯛魚隻充分互動,交流基因,加上養殖真鯛每一個世代其生存率減少約 12%的天擇壓力被淘汰,而逐漸去除與漂白鹿兒島灣內部養殖魚基因的比率而降低占比。
1998年~瀨戶內海的鰆魚栽培漁業
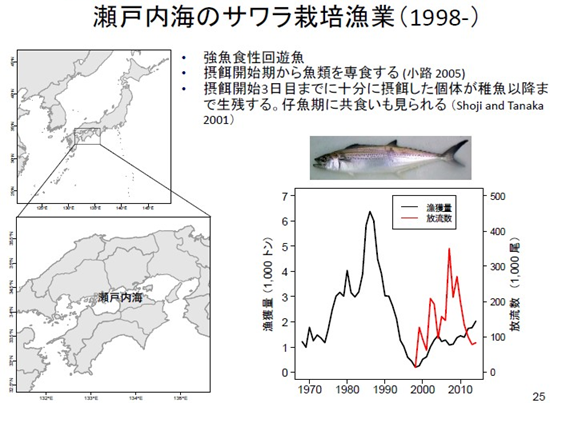
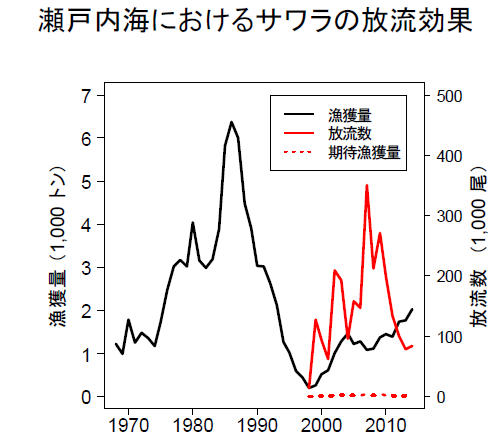
日本瀨戶內海 1980 年代原本鰆魚產量很高,可能過度撈捕,80 年代中期後漁獲急速下降(右圖黑線),日本於 1998 年開始放流鰆魚魚苗(右圖紅線)。
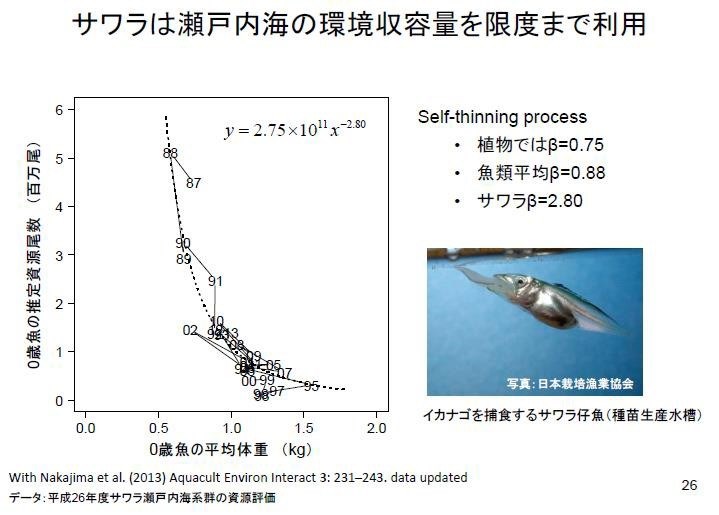
單位體積的海域之內,放流鰆魚魚苗越多,鰆魚0歲魚的平均體重越小,也即成長的越慢,呈負相關。呈現自我稀疏(Self-thinning process):意指在一定生態環境承載量,個體數一旦增加,則每個個體的體重減小現象。日本研究發現鰆魚是洄游性的獵(掠)食性魚類,小魚會同類互噬(cannibalism)、β 指數為 2.80,是一般平均魚類(0.88)β 值的 3.18 倍,植物的β值僅 0.75。
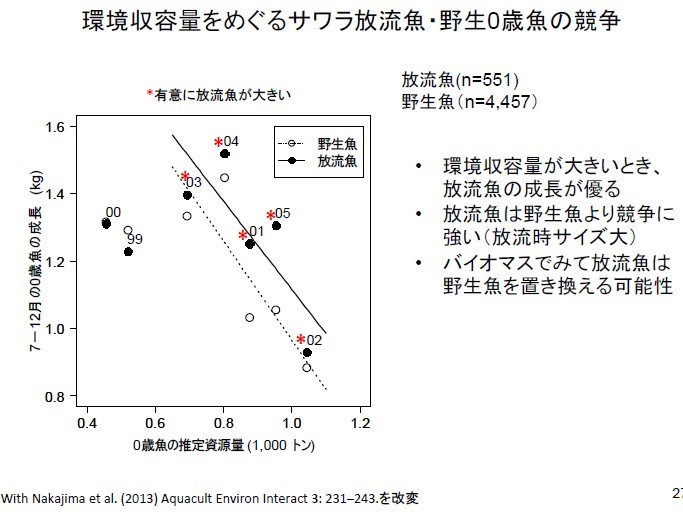
日本研究統計在瀨戶內海放流 6 年(1999~2005 年)鰆魚的資料,發現當環境承載量很好的時候(2001~2005 年),養殖鰆魚的 0 歲放流魚苗(同一年度垂直線上的黑圓圈)會顯著(*)長的比野生的(同一年度垂直線上的白圓圈)大。因為養殖鰆魚放流魚苗放流時的魚體較大,因此會比野生鰆魚魚苗更具競爭優勢,而且因為瀨戶內海環境可以提供的生物資源(biomass)是一定的,所以放流養殖魚可能會取代野生魚。
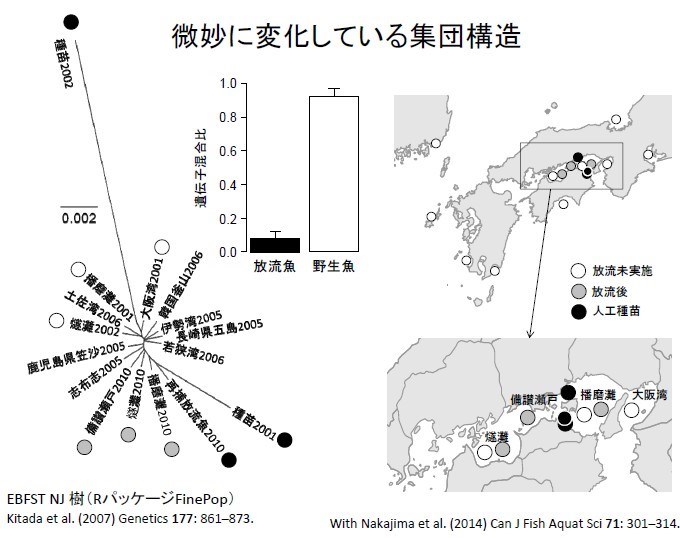
日本曾經在瀨戶內海用 10 尾捕獲野生的鰆魚種魚,培育 10 萬尾魚苗放流,研究基因是否會對瀨戶內海造成生態的影響。研究發現,放流並沒有影響瀨戶內海鰆魚族群遺傳的多樣性。但是鰆魚族群的遺傳距離起微妙變化。
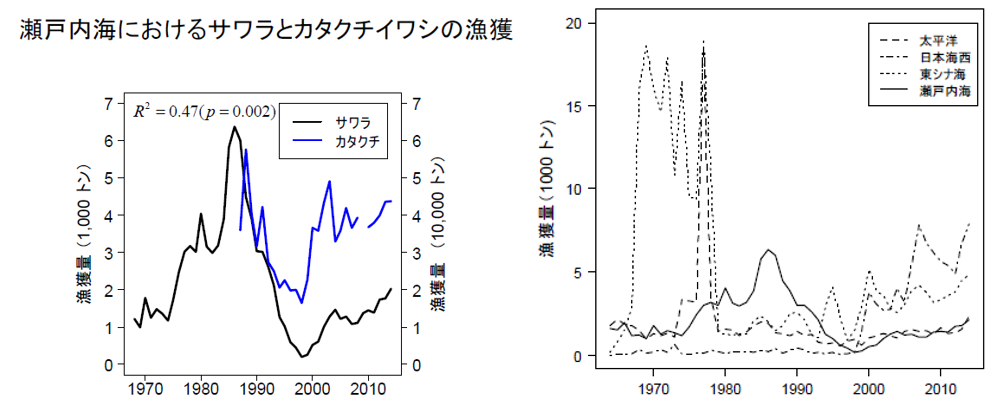
日本只在瀨戶內海(實線)放流鰆魚(左上圖)。因為同時期日本其他沒有放流鰆魚(虛線)的海域(日本海、日本海西、東海)的鰆魚數量也是增加(右上圖),所以海域鰆魚漁獲量的增加,應該是與海域的食物來源增加有關,例如鯷魚(カダクチイワシ:左上圖藍線);而和是否有放流鰆魚魚苗無關。因為日本計算瀨戶內海(下圖黑實線)放流鰆魚的YPR 值(放流後野外捕回重量;紅點線), 發現鰆魚放流量只佔總漁獲量的 2.5%貢獻度(下圖);反而鯷魚(藍線)的貢獻度47%(左上圖),占近 5 成 (統計相關達 R2=47%)。所以瀨戶內海海域鰆魚漁獲量的增加,明顯是與海域的鯷魚食物來源增加有關。
綜整與結論:
1. 放流的數量跟自然入添量的變動相比,顯然微不足道,所以很難確定放流是否有效果,而進行栽培漁業應以一代再捕利用回收為原則。
2. 放流在第一代的直接效果,是靠著種苗活存率還有成長率來決定,成功放流意味可以回本。由於環境承載量固定,放流魚與野生魚產生競爭,甚至養殖放流魚會取代自然界的野生魚,卻沒有增加自然界的生物量。
3. 人工養殖海水魚的基因跟野生海水魚的相差很多,但因為養殖魚在野外的存活率比較低,加上天擇作用與養殖族群和野生族群的交配,放流養殖魚的基因會逐漸從野生魚群的基因中被淘汰掉。
4. 漁業資源的回復,應該是要保護魚類的產卵場、修復仔魚孵化育成場的棲地;保護棲地比放流水產生物種苗有效。
檢討與未來建議:
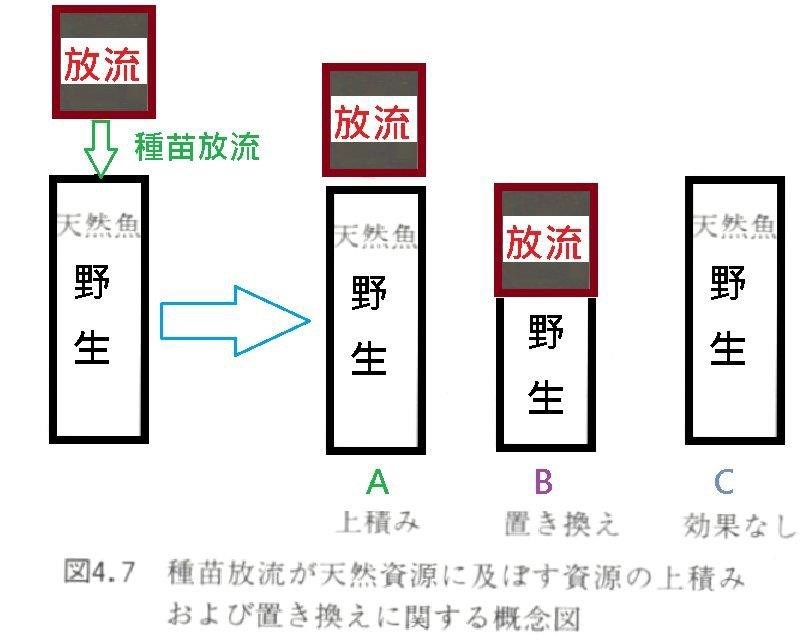
1. 環境的生物承載量是一定的。因為養殖放流種苗與野生種苗劇烈的競爭結果, 只會發生養殖放流魚苗取代野生魚隻的作用,所以不過是生物取代、置換而 已,總生物量並沒有增加。北田教授研究也顯示放流種苗雖不影響野生族群基因 的多樣性,但放流種苗長大顯然與野生魚種產生交配繁衍下一代,若以遺傳 距離定義野生種與放流種,兩者遺傳結構仍有差異產生。
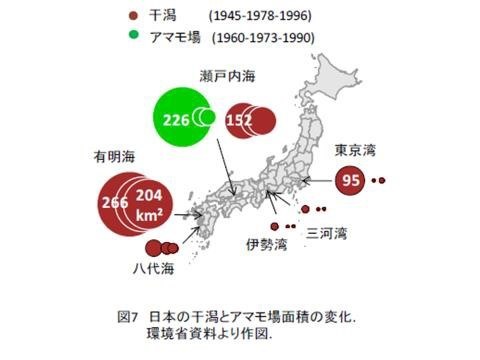
2. 從鰆魚不必放流,資源量也成長的結果來看,據日本環境廳的資料顯示,日本潟湖(干潟)和藻場(アマモ場)和沒工業化前相較已減少很多,重工業化地區如東京灣潟湖和藻場減少更多。與其花錢與資源在放流種苗,應該要著手於棲地的復育。
3. 愈來愈多的研究顯示,無意識的水產養殖作業方式及不當的人工魚苗放流, 反而會造成生態失衡。隨著分子標記技術的進步、價格平價與普及化,已證實不當人工放流水產養殖生產的產品,反而造成污染天然族群之基因、混雜品系、引進入侵種及病源菌等反效果,嚴重破壞及攪亂原本物種的遺傳結構, 造成族群弱化,生態失衡,加速自然資源的枯竭。
4. 在此次演講的最後,北田教授也再度誠摯的呼籲,放流的必要性應作審慎評估,對魚類養殖初期生態需更深入了解,個體回復後更應針對該物種的產卵場、孵育場進行保育,配合放流政策再生產。
5. 種苗放流的必要仍是值得討論的議題,放流對於其他物種之影響,養殖魚種與野生種之基因交流,皆是未來研究應該可著力的方向,希望往後能找到最適當的物種保育方式,不僅讓海洋資源可以永續的發展,於經濟效益上也更划算。
對比臺灣漁業水產種苗放流的現況與建議
雖然臺灣漁政縣市政府單位每年均有編列巨額經費進行養殖種苗人工放流,但至今仍有許多需要未周全考慮之遺傳與環境因子,如:人工繁殖魚種與野生魚種有基因之差異,是否會有污染天然族群遺傳基因,破壞棲地或生態失衡的問題。此外,臺灣漁政公營單位多不生產種苗,臺灣在進行種苗人工放流時,往往是由數個私人養殖場競標,由得標者提供漁政公營單位放流所需的種苗數量。在種魚來源及遺傳資訊不明,魚苗難以分辨(魚苗辨識不易、放流魚苗是否是當地缺少的物種等隱蔽物種問題),只粗略檢查兩種(神經壞死和虹彩)病毒反應(完全忽略細菌、真菌、寄生蟲等其他微生物),及 4 種(含氯黴素、孔雀綠、還原型孔雀 綠 、 硝 基 呋 喃 代 謝 物 ) 藥 物 殘 留等,在品質不清的情況,以及缺乏族群遺傳及生物多樣性的概念下,往往使得放流效果事倍而功半,無法確實達到預期之效益。
同時「中間育成」技術也是決定種苗放流成功與否的重要關鍵。日本很慎重的把未來要放流的稚魚種苗,由陸上室內養殖場移送到設置於海上或河川之戶外箱網或圍網,讓養殖的稚魚種苗接受嚴酷考驗數十日不等,以增強其野放後的警戒心與競爭能力,為適應未來的大自然環境預先做準備。我研究室解剖了台灣放流的養殖黑鯛與野生黑鯛比較,發現野生黑鯛的肝是正常的紅色,養殖黑鯛竟然是粉紅泛白色,推測可能是吃了太多飼料又因為密集養殖缺乏運動變成了脂肪肝。再以日本栽培漁業生產放流用的嘉鱲種苗為例,野生的嘉鱲魚體色是紅色的,人工養殖嘉鱲魚因為長期人工養殖,生長環境與食物不同,竟然變成黑色無光澤,甚至連鼻孔,從兩個變成了一個。日本也曾經研究魚苗放流後的沉底速度,發現野生的魚苗警戒性高,會快速沉底尋找掩蔽,反之人工養殖魚苗會停在水層表面,導致被捕食的機率大大增加。因此沒有完善的中間育成訓練,貿然放流只讓這些魚去送死。
直到將被放流的種苗之視覺、嗅覺感官,調節滲透壓等生理機能、與游泳或潛砂能力等型態上的運動器官發育至某程度、逐漸能夠主動自己覓食、學習調整行為,例如懂得逃避敵害,發展出具備野外求生的基本能力;亦即,由人為選擇轉換為天然選擇,先淘汰不適應野外環境的種苗後,去蕪存菁再放流,以提高放流後的種苗活存率。然而在臺灣,針對放流魚種之「中間育成」步驟較為缺乏,大多為在種苗放流前和運輸過程中短暫馴化,得標廠商在種苗放流前 3 日,將種苗易池到底質非泥土之容器,注入現喞海水蓄養 3 日。期間並停止餵食魚苗,使魚苗腸道淨空,降低排洩物產生,減少運輸時的水質污染,提高魚苗活存率,也就是把至今為止一直生活習慣於養殖池水質條件的養殖魚苗,瞬間轉置於天然海水鹽度,迫使其緊迫適應 3 日,使得放流到天然海域後的養殖魚苗存活率普遍不高。其次盡管台灣沿岸就有六種黑鯛物種,南部與北部沿岸黑鯛可能是完全不同物種,若是採用招標採購常常變成南魚北運, 養殖魚苗被瞬間放流至魚種、水文氣象與海洋環境完全陌生的場域。此外台灣除了少數魚種及少量樣本曾進行標識試驗研究,例如估計黑鯛魚苗年度放流貢獻度在 1.7%~6.6%之間、但絕大部分放流的水產生物並未標識,未進行放流效果之評估。而傳統的標記方式(生物、物理、化學)更無法估計放流水產生物的繁殖率、死亡率及再生產量。使得臺灣水產研究者無法進行完整周延的放流評估,人工放流的成效和對天然生態的影響(甚至造成反效果)實在堪慮。
另外,民間盛行宗教放生以積功德,往往引起許多生態保育與動物保護人士團體的撻伐。的主要原因在於:(1) 商業化的放生事實上只是鼓勵了額外供給食用魚市場的繁養殖行為,並未如宗教團體所預期的能護生;(2) 在放生過程中因為搬運所產生的動物緊迫與傷害一直不被佛教界所看見,放流的程序粗糙, 最後造成放生等於放死,而這樣的後果又被解讀為捨報。此外,(3) 大量放生動物最令人垢病的環境生態衝擊除了在瞬間於環境中投入大量動物所增加的環境負擔、造成入侵種問題、造成原生生物群聚的改變之外,最 糟糕的便是對長期以來經由海潮、地質變動與生物演化自然而然所形成的遺傳與 物種多樣性,而擾亂這樣的多樣性,事實上正與永續利用與保育漁業資源是背道而馳的。
今天的農業與畜牧的大多數動物和植物,牠和它們已和野生祖先在遺傳上有相當程度的不同,例如牠和它們需要人類的照料、呵護和提供食物飼養以維持生存,例如:牛,羊,豬、雞,火雞,很少還保有牠們祖先型的野性,然而在水產養殖中使用的物種大多仍是野生族群,尚未完全被人為地飼養馴化,而且大多數如果釋放或逃脫返回原先棲地仍有能力恢復野性,放流養殖魚苗和逃脫的水產養殖品種,經常傳染疾病與媒介引入寄生蟲給野生物種也時而耳聞,例如逃逸的大西洋鮭魚身上的魚蝨嚴重感染到野生的大西洋鮭魚,魚苗放流是否也有類似的隱憂值得深慮。
在浩瀚的海洋耕耘水產養殖,把海洋當牧場的利弊必須評估其對生態等其他難以計量的成本,例如文化和景觀。首先最應該考量的基本問題是:孵化場是否實際上在魚隻的生產中具有成本效益,以及目前孵化場的實際操作是否實質上增加了所欲增產魚類的生物量,還是只是以孵化場生產的魚隻取代野生魚隻,甚至在取代的過程中甚至與野生魚類競爭棲地等,均應該與所產生的環境成本進行權衡,同時絕對不應忽略恢復棲息地和調控監管撈捕漁獲的功能。
根據日本累積 60 年的經驗,水產種苗放流的成功與否還必須借助:(1) 放流後的種苗有適當的棲地,以供覓食與逃避敵害,而一般大多數的海洋魚類及水產無脊椎動物需要一種以上的棲地;豐富充足的食物與良好的天然環境來完成其生活史;(2) 立法規定漁獲體長、限制漁法、捕獲季節等人為劍及履及的執法, 積極的資源自主管理。雙管齊下,栽培漁業才有望大功告成。
參考文獻
1. 北田修一 (2016)。種苗放流の効果と野生集団への影響。日本水産学会誌。82(3) 241-250。
2. 北田修一 (2014)。遺伝標識による種苗放流効果の推測:現状と適用上の留意点。日本水産学会誌。80(6) 890-899。
3. 北田修一 (2013)。シロザケ孵化場魚の遺伝的多様性と繁殖成功度. 「サケ学大全(帰山雅秀,永田光博,中川大介」編)」, pp. 123-128, 北海道大学出版会, 札幌).
4. 北田修一 (2012)。標識放流. 「最新水産ハンドブック」, 講談社サイエンティフィク.
5. 北田修一 (2011)。種苗放流の遺伝的影響:実態と展望. シリーズ 21 世紀の農学
6. 「農林水産業を支える生物多様性の評価と課題(日本農学会編)」, pp. 83-112,養賢堂, 東京.
7. 北田修一 (2010)。栽培漁業の遺伝的影響に関する実証的考察. 平成 21 年度栽培漁業技術中央研修会テキスト集, 水産総合研究センター, 14p.
8. 北田修一 (2008)。種苗放流の遺伝的影響. 水産資源の増殖と保全, pp. 190-213,成山堂書店, 東京.
9. 小畑泰弘, 山崎英樹, 岩本明雄, 浜崎活幸, 北田修一 (2008)。環境収容力と種苗放流―瀬戸内海東部海域におけるサワラの例. 水産資源の増殖と保全, pp. 66-85,成山堂書店, 東京.
10. 浜崎活幸, 北田修一 (2008)。環境変動と資源増殖―アワビ類を事例として. 水産資源の増殖と保全, pp. 86-104, 成山堂書店, 東京.
11. 北田修一 (2007)。沿岸水産資源の増殖と管理. 農林水産技術研究ジャーナル 30 (12), 20-25.
12. Kitada, S. (2018). Economic, ecological and genetic impacts of marine stock enhancement and sea ranching: A systematic review. Fish and Fisheries. 1–22.
-

-
郭金泉
教授
國立海洋大學水產養殖系
關鍵字
文章摘譯
